Why Dynamic Single-Molecule?
The complexity of transcription mechanisms
DNA transcription is the key element that defines cellular identity and status. By clearly understanding the mechanism of gene regulation and expression at this level scientists will be able to grasp many causes of diseases and develop possible cures. Until now, molecular biology and biochemistry methods have highly contributed to uncovering the transcription mechanism. Without looking at the dynamics of the individual components in real-time and at the molecular level, it will not be possible to fully comprehend this complex process.
Overcome these challenges with Dynamic Single-Molecule technology through:
- Visualize single DNA polymerase in real-time
- Base-pair level observation of DNA transcription dynamics
- Observe while manipulate the transcription process at the same time
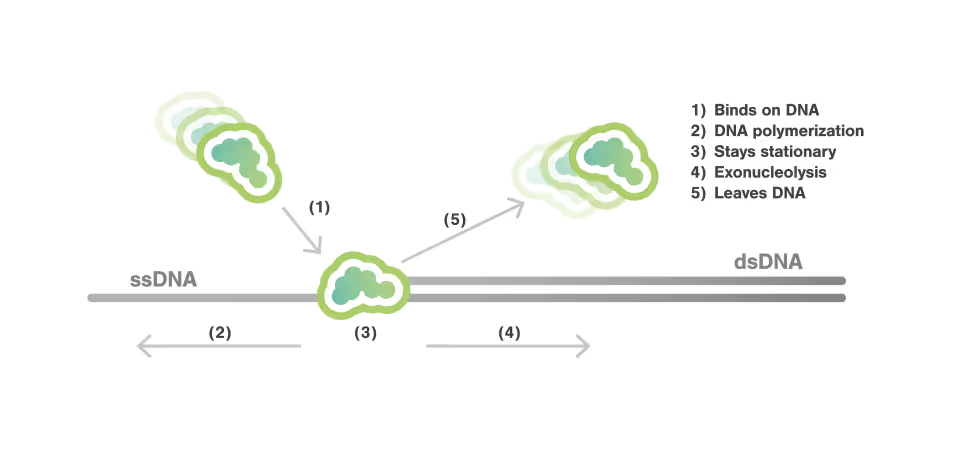
Real-time observation of DNA exonuclease dynamics at base-pair level

In this experiment, the exonuclease activity of the T7 polymerase was investigated. This is a DNA polymerase from the T7 bacteriophage that copies DNA strands in the 5’→ 3′ direction, and also features exonuclease activity. For this analysis, an optically trapped double-stranded DNA was held at a constant force that induced exonucleolytic activity of the polymerase. By removing nucleotide after nucleotide from one strand, the polymerase was unwinding the dsDNA. As the length of single-stranded DNA is longer compared to its double-stranded state, the unwinding resulted in a gradual increase in the end-to-end distance of the DNA (Figure a). This change in length was directly translated into the activity of the T7 DNA polymerase and the number of nucleotides it processed over time (Figure b). Specifically, short activity bursts ranging between 3 and 10 nucleotides were revealed, interspersed by frequent pauses of varying duration. This provided deeper insights into the dynamics of T7 exonuclease activity.
Figure a Force-distance curve of double-stranded DNA (blue) and single-stranded DNA (red).
Figure b Activity bursts of T7 DNA polymerase performing force-induced exonucleolysis on a double-stranded DNA.
Dive into the publication
Solutions
C-Trap
Molecular biology as never seen before
The C-Trap® provides the world’s first dynamic single-molecule microscope to allow simultaneous manipulation and visualization of single-molecule interactions in real time.

Technical note:
These cards are NOT components because they use the finsweet nested collection logic. To pull in the posible multiple people wo worked on it.
This type of 1-many relation is not supported native in Webflow.
Also this section is hidden when emtpy. To keep everything visible here, that is being done outside the webflow designer from within Slater.
These cards are NOT components because they use the finsweet nested collection logic. To pull in the posible multiple people wo worked on it.
This type of 1-many relation is not supported native in Webflow.
Also this section is hidden when emtpy. To keep everything visible here, that is being done outside the webflow designer from within Slater.
Kinetic control of mammalian transcription elongation
Wang, Y. et al.
2025
Nature Structural & Molecular Biology
This is some text inside of a div block.
Phase-separated NDF−FACT condensates facilitate transcription elongation on chromatin
Li, Z. et al.
2025
Nature Cell Biology
This is some text inside of a div block.
Chromatin sequesters pioneer transcription factor Sox2 from exerting force on DNA
Nguyen, T. et al.
2022
Nature Communications
This is some text inside of a div block.
.avif)




SITC 2025
Conference
April 22, 2025
01-01-20
This is some text inside of a div block.
CAR-TCR Summit 2025
Conference
April 22, 2025
01-01-20
This is some text inside of a div block.
CICON 2025
Conference
April 22, 2025
01-01-20
This is some text inside of a div block.